Redox-regulation and interactome of the stress-sensing chaperone Get3
Oxidative stress can cause ATP depletion, oxidative protein unfolding and potentially toxic protein aggregation, which endangers the integrity of the entire cell and has been associated with many diseases and aging (1). The drop of ATP levels poses a particular challenge, since cells usually rely on ATP-dependent systems to counteract protein unfolding. Therefore, the activation of ATP-independent chaperones "holdase", which bind unfolding protein intermediates to prevent their aggregation, serves as first line of defense (2). The highly conserved, cytosolic yeast protein Get3 (TRC40/Asna1 in mammals) switches from its ATP-dependent targeting function for tail-anchored membrane proteins into a general, ATP-independent chaperone protecting proteins from aggregation upon oxidative unfolding stress (Figure 1) (3, 4).
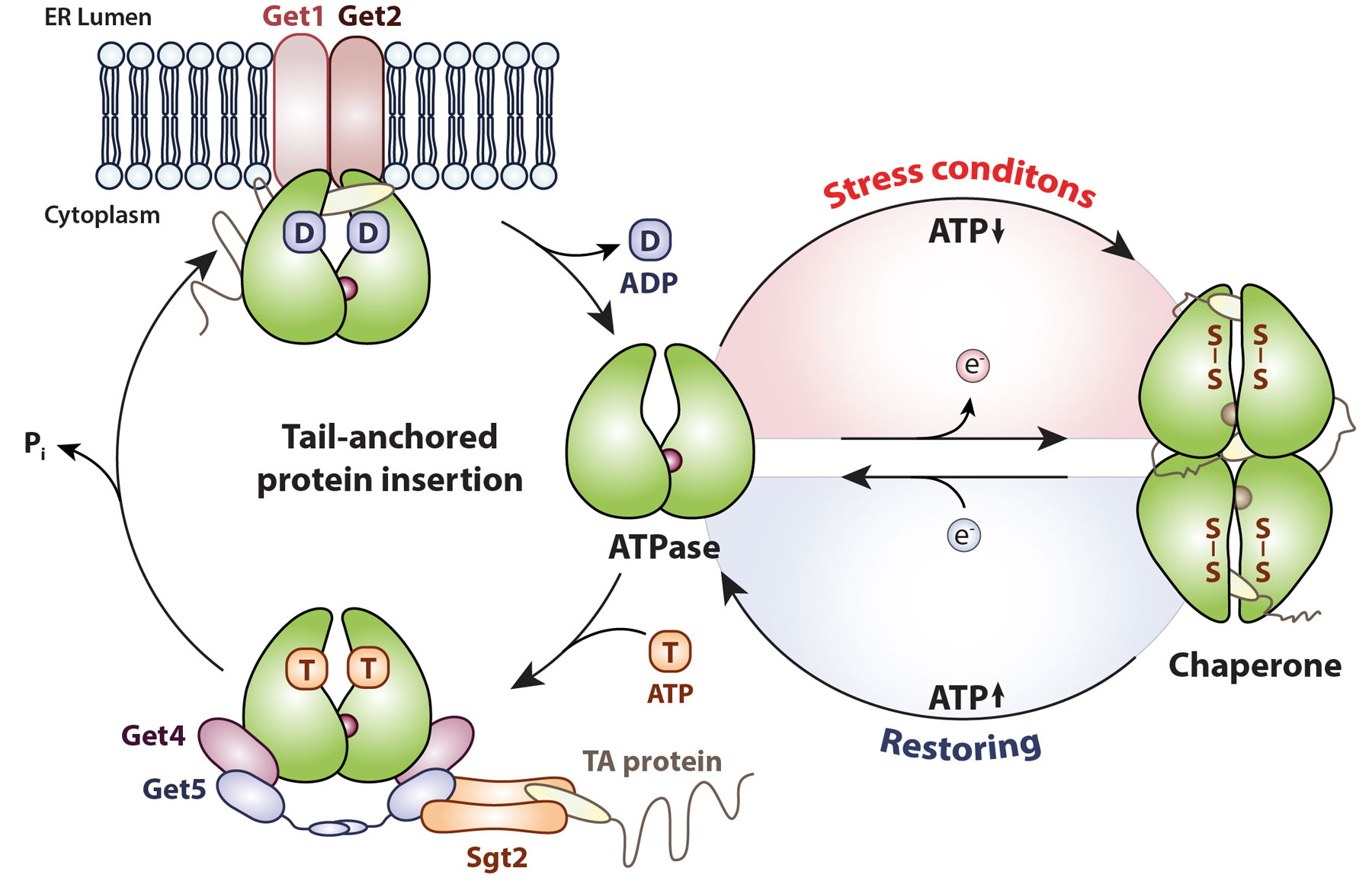
Under non-stress conditions, Get3 functions as an ATP-dependent targeting factor in the GET pathway mediating the Guided Entry of Tail-anchored (TA) proteins into the ER membrane. ATP-depleting, oxidative stress conditions cause disulfide bond formation and major conformational changes turning Get3 into an ATP-independent chaperone. The switch is fully reversible upon restoring non-stress conditions. Figure modified from Ulrich et al. 2020
Our lab recently showed that Get3’s functional switch depends on the redox state of the highly conserved CXC motif which is under tight control of Get3’s nucleotide-binding state, making Get3 an ideal sensor for ATP-depleting, oxidative stress conditions (5). Nucleotide release allows the redox-sensitive cysteines to react with oxidants, such as H2O2 and GSSG. Subsequent disulfide bond formation causes the partial unfolding and formation of chaperone-active oligomers. Vice versa, inactivation of the chaperone function depends on the initial reduction of Get3 followed by rebinding of ATP. This two-step mechanism ensures that clients are only released upon restoring both, the cellular redox homeostasis and ATP-levels, which are required for the energy-driven proteostasis system to handle unfolded clients. Lack of the redox-sensitive cysteines impairs the transfer of client proteins to the ATP-dependent Hsp70/Hsp40 system upon stress recovery and renders yeast cells more sensitive towards H2O2 stress (5).
We now aim to identify clients that depend on Get3’s stress-induced chaperone function and focus on the crosstalk between the cellular redox system and proteostasis during acute stress conditions and recovery. Using strains that lack the glutathione or thioredoxin system, we test which cellular redox system mediates the reductive inactivation of Get3’s chaperone function upon stress removal. We further study the crosstalk of redox enzymes with the cellular proteostasis system to understand the fate of released clients focusing on refolding as well as potential proteasomal and lysosomal degradation.
1. Reichmann D, et al. (2018) Maintaining a healthy proteome during oxidative stress. Mol Cell, 69(2):203-13. doi:10.1016/j.molcel.2017.12.021
2. Ulrich K, Schwappach B, Jakob U. (2020) Thiol-based switching mechanisms of stress-sensing chaperones. Biol Chem. doi:10.1515/hsz-2020-0262
3. Powis K, et al. (2013) Get3 is a holdase chaperone and moves to deposition sites for aggregated proteins when membrane targeting is blocked. J Cell Sci. 2013;126(Pt 2):473-83. doi:10.1242/jcs.112151
4. Voth W, et al. (2014) The protein targeting factor Get3 functions as ATP-independent chaperone under oxidative stress conditions. Mol Cell, 56(1):116-27. doi:10.1016/j.molcel.2014.08.017
5. Ulrich K, et al. (2022) From guide to guard-activation mechanism of the stress-sensing chaperone Get3. Mol Cell,82(17):3226-38 e7. doi:10.1016/j.molcel.2022.06.015